




Содержание
Перейти к:
https://doi.org/10.32454/0016-7762-2020-63-1-54-62
Перейти к:
Кузнецов В.Г. АСИНХРОННОСТЬ РАЗВИТИЯ РИФОВ И РИФОСТРОЯЩЕЙ БИОТЫ. ПАЛЕОЗОЙ. Proceedings of Higher Educational Establishments: Geology and Exploration. 2020;1(1):54-62. https://doi.org/10.32454/0016-7762-2020-63-1-54-62
Kuznetsov V.G. ASYNCHRONOUS DEVELOPMENT OF REEFS AND REEF-BUILDING BIOTA. PALEOZOIC. Proceedings of higher educational establishments. Geology and Exploration. 2020;1(1):54-62. (In Russ.) https://doi.org/10.32454/0016-7762-2020-63-1-54-62
Постановка задачи и объект исследования
В сферу интересов европейской науки рифы вошли в эпоху великих географических путешествий и открытий и более осознано и целеустремленно — с конца XVIII века, когда во время второй экспедиции Дж. Кука был открыт Большой Барьерный риф Австралии, а участник этой экспедиции немецкий натуралист Рейндгольд Форстер установил, что эти рифы созданы кораллами, и возникло представление о коралловых и, в более общем виде, органогенных рифах. Уже в начале XIX века были выявлены древние ископаемые рифы. В первой половине XX века стало ясно, что органогенные сооружения достаточно разнообразны, были введены понятия «биостром» и «биогерм», а во второй его половине — «органогенная постройка» (organic buildup). Последний термин, по остроумному замечанию академика О.С. Вялова, оказался «безалкогольным», то есть крайне широким и неопределенным. Были предприняты попытки создать некоторую иерархию: первый уровень — органогенная постройка, которая включала биостромы и биогермы, и второй, более высокий — собственно рифы. При этом в определение кроме механизма образования — за счет жизнедеятельности организмов — включался и второй показатель — морфология постройки: пластовая в случае биострома и холмовидная в случае биогермов и рифов.
В свою очередь, холмовидные образования начали подразделяться по наборам строящих их организмов. Так, В. Кисслинг и Э. Флюгель [20] подразделили подобные сооружения на три типа:
Обзор основных терминов, относящихся к обсуждаемой тематике, был проведен в специально подготовленном словаре-справочнике, где риф определен как «сложное геологическое образование, возникшее в результате жизнедеятельности колониальных или нарастающих организмов, живших вблизи поверхности воды и обладающих волноустойчивым потенциалом и представляющее собой карбонатный массив, сложенный, по крайней мере частично, остатками организмов и продуктами их разрушения, возвышающийся в период своего формирования над дном окружающего моря и образующий волнолом. Поскольку скорость роста рифа превышает скорость накопления окружающих осадков, его мощность больше мощности синхронных отложений» [14].
По сути дела, это определение включает два первых типа построек В. Кисслинга и Э. Флюгеля. В аналогичном значении понятие «иловый холм» используется и в настоящей работе.
Мировая литература по рифам, их биоте, составу, строении, распространении в пространстве и времени огромна. В 1992 г. Э Флюгель и Э. Флюгел-Калер составили библиографию по фанерозойским рифам, которая заняла более 100 журнальных страниц формата А4 и включала более
5000 наименований [17]; количество публикаций по проблеме с тех пор существенно возросло, причем как в нашей стране, так и за рубежом были изданы монографии и специальные тематические сборники [3, 12, 13, 22, 23 и др.] и многие сотни статей. Другими словами, материал по ископаемым рифам огромен, и создавать еще какие-либо обзоры по распределению рифов по площади и в стратиграфическом разрезе, равно как и подробно характеризовать рифы разных возрастов и регионов, в целом нет никакой необходимости, поэтому в данной работе приведен лишь краткий обзор и краткая характеристика рифов палеозоя.
Для решения поставленной задачи требуется очень дробная стратиграфия и точная корреляция узких стратиграфических подразделений, в том числе между весьма удаленными друг от друга регионами, что по ряду причин удается далеко не всегда, но в целом общую картину выяснить более или менее достоверно, с теми или иными затруднениями, все же возможно, что и составило содержание настоящей работы.
Распределение рифов в разрезе палеозоя
В палеозойской истории установлено пять крупных циклов рифообразования: раннекембрийский, ордовикский, самый продолжительный и самый значительный по количеству регионов их распространения и мощности отдельных сооружений, силурийско-франский, поздневизейско-серпуховский и пермский, верхние границы которых определяются моментами массовых вымираний на границах ранний-средний кембрий, ордовик-силур, фран-фамен, серпухов-башкир и пермь-триас (рис.).
Характеристика рифов соответствующих этапов и их биоты дана в серии предыдущих публикаций [2—9 и др.], поэтому здесь не рассматривается, а обсуждаются более конкретные вопросы соотношения собственно рифообразования как такового и рифостроящей биоты.
Кембрийский цикл рифообразования был относительно кратким в палеозойской истории Земли — рифы развивалось в течение 18—19 млн лет.
Рифы этого цикла развиты в Алтай-Саянской складчатой области, в Монголии, Канаде, Австралии и на Сибирской платформе, где они, по-видимому, наиболее изучены. Каркасообразующими организмами были археоциаты; аналогичные функции, возможно хотя бы частично, выполняли кустистые эпифитоны. Основной объем карбонатного материала поставляли водоросли и микробиальные сообщества.
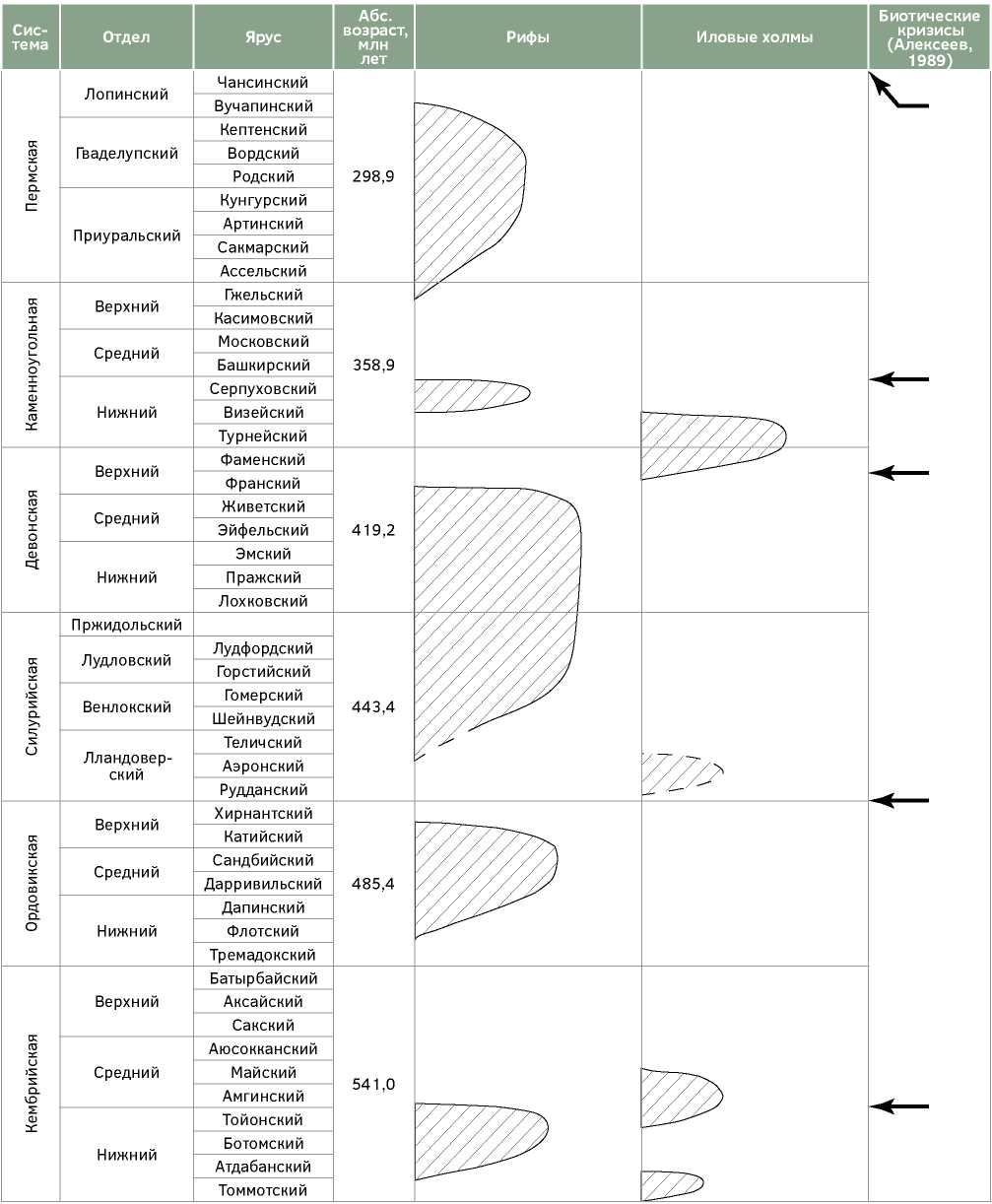
Рис. Схема цикличности рифообразования палеозоя
Fig. Scheme of cyclicity of Paleozoic reef formation
Дробная стратиграфия отдельных ярусов нижнего кембрия, которая, как отмечено выше, нужна для целей настоящего исследования, практически отсутствует, однако некоторые примеры рифов Сибирской платформы, где установлены самые древние рифы, позволяют предполагать, что собственно рифы начинают формироваться не с самого начала кембрия — в основании осинского горизонта вначале отлагаются слоистые карбонатные отложения, в том числе и с археоциатами, и лишь затем начинают формироваться собственно рифовые сооружения.
После первого фанерозойского вымирания на границе тойонского и амгинского ярусов рифообразование прекратилось, возникали постройки только микробиально-водорослевые, а возможные каркасообразователи в виде губок развивались вне рифового биоценоза.
Средний, поздний кембрий, ранний и начало среднего ордовика — тремадокский и аренигский века общей стратиграфической шкалы, тремадокский, флосский, дапинский и начало дарривилского веков международной шкалы — это время отсутствия рифов, становление которых относится лишь во второй половине дарривилского века (лландоверийского общей шкалы). Важно отметить, что каркасные организмы — потенциальные рифостроители появились в геологической истории раньше — строматопороидей и кораллы в тремадоке и раннем арениге, мшанки в конце раннего — начале среднего ордовика [12, 24 и др.].
Другими словами, рифовый биоценоз, который и привел к формированию морфологически выраженных «истинных рифов», сложился позднее возникновения собственно составляющих его организмов.
Основное ордовикское рифообразование с некоторыми колебаниями его интенсивности продолжалось порядка 24—25 млн лет и происходило в дарривилском, сандбийском и катийском веках, и в ограниченных масштабах в начале хирнантского века, хотя каркасные возможно рифостроящие организмы еще существовали [5].
После мощного, но в целом кратковременного и не столь катастрофического, как следующее фран-фаменское, вымирания на границе ордовик-силур начался третий, самый длительный — порядка 69 млн лет силурийско-девонский цикл рифообразования, который закончился до конца девона — на границе франа и фамена. Он также является периодом и самого масштабного рифообразования как по числу областей развития рифов, так и по мощностям и разнообразию построек. Этот цикл состоит из двух этапов с относительным сокращением формирования рифов в начале девона — лохковском и пражском веках, что, однако, не повлияло принципиально на состав основных рифостроителей, которыми, как и ранее, были строматопороидеи, кораллы, главным образом табуляты, и мшанки. Закончился этот цикл на стратиграфической границе относительно низкого уровня — франского и фаменского веков.
Распространены рифы силурийско-франского этапа в пределах Восточно-Европейской (Русской) платформы и Урала, Северо-Американской платформы и Кордильер, в Казахстане, Алтай-Саянской и Верхояно-Колымской областях, на Чукотке, в Монголии, Китае и Австралии, то есть практически по всему земному шару [3, 4, 7—9, 1 2, 23 и др.].
Ряд небольших построек, часто даже не соответствующих принятому в настоящей работе понятию рифа, описан в начале силура — в нижнем руддании, в частности в бассейне Мичиган [15, 16], однако дальнейшего развития они не получили и силурийское собственно рифообразование началось лишь в конце лландовери, а массовое с венлока, возможно, скорее, даже не с самого его начала.
Основными организмами-каркасообразователями силурийско-девонского этапа были кораллы, главным образом табуляты, строматопороидеи, мшанки. Как и всегда в рифах, обильны водоросли и микробиальные сообщества как важные поставщики материала и его цементации.
Что касается окончания рифостроения, то на ряде конкретных примеров востока Русской платформы, Западно-Канадского бассейна, Австралии, Китая и горно-складчатых областей (Гарц, Германия) показано, что собственно рифы исчезли ниже границы фран-фамен, до события Кельвассер, которое завершило вымирание, когда потенциальные рифостроители еще существовали, но вне рифового биоценоза [4—9].
После массового вымирания на границе франафамена каркасное рифостроенние в рассматриваемом в данной работе смысле практически прекратилось, шло формирование иловых холмов в их «классическом» выражении, в частности в виде уолсортских фаций. В ряде случаев в них встречаются остатки и каркасных рифостроителей, например в фаменских сооружениях Печорской синеклизы в виде разрозненных строматопороидей, но каркасообразующего значения они не имели. Временной интервал подобного развития иловых холмов: фамен — средний визе.
С позднего визе каркасное рифостроение возобновилось, и оно охватило серпуховский век. Это был самый короткий в палеозое цикл рифообразования, который продолжался порядка 16 млн лет.
Рифам этого цикла посвящена специальная статья [6], поэтому можно ограничиться очень кратким описанием этих образований. В пределах Евразии они известны в Западной Европе в Великобритании, Бельгии и Франции, в Донбассе, на западных и восточных склонах Урала, в Прикаспийской впадине по ее западному обрамлению и непосредственно во впадине в виде крупных массивов Тенгиз, Карачаганак и др., в Восточном Казахстане, на северо-востоке в Алазейской складчатой области и Сихотэ-Алине. Рифы этого возраста описаны на востоке Канады и США, в Австралии. Каркасными строителями были мшанки и кораллы (преимущественно табуляты), но везде в значительных, а иногда и преобладающих (Злоказовские рифы Западного Урала) присутствовали микробиально-водорослевые компоненты.
Не самое значительное по масштабу вымирание организмов на границе серпухов-башкир определило, однако, длительное — в течение среднего и позднего карбона, прекращение рифообразования, хотя потенциальные рифостроители в виде кораллов, мшанок и строматопорат еще существовали. Следует, однако, отметить, что количество последних было существенно ограничено, равно как и кораллов, разнообразие которых, по данным А.С. Алексеева [1], на этом рубеже сократилось на 80%.
К сожалению, не удалось найти разрезы с очень дробными и детальными подразделениями, поэтому не установлено, прекратилось ли рифообразование точно на рубеже вымирания на границе серпухов-башкир, или оно завершилось несколько раньше, как это установлено в конце ордовика и конце франа. Что касается начала рифообразования, то оно, как отмечено выше, произошло значительно позднее франско-фаменского вымирания.
Новый, последний цикл палеозойского рифообразования датируется пермским периодом и продолжался, видимо, 45—47 млн лет.
Нижне- и среднепермские образования широко развиты на Русской платформе, в Прикаспийской впадине, на Урале, на Северо-Американском континенте, в пределах Северной и Центральной Европы, средне-верхнепермские на Кавказе, Памире, в Иране, Пакистане, Китае, где они, по-видимому, наиболее подробно изучены.
Состав пермских рифостроителей существенно отличается от такового предыдущих этапов. Практически потеряли свое значение кораллы, в ранней перми относительно сохранили эти функции мшанки, но основная роль перешла к губкам, разнообразие таксонов которых, особенно в поздней перми, существенно расширилось. Для целей настоящей работы очень важные сведения предоставляют рифы Южного Китая, где показано, что собственно рифы ни в одном случае не достигают границы перми и триаса, разрезы самого конца перми над рифами завершаются карбонатными слоистыми отложениями с тем же набором организмов, что и рифовые образования [11,21].
Соотношение рифообразования и развития рифостроящих организмов
Приведенный краткий обзор временного развития рифов и рифостроящих организмов показывает отсутствие строгого соответствия между ними.
Оставляя пока кембрийский цикл, следует отметить, что после появления рифостроящей биоты необходимо определенное время, чтобы эти организмы сформировали определенный биоценоз, определенную экосистему, которая уже и создает рифовые постройки. Это отчетливо видно на примере ордовика, когда соответствующие организмы возникли в раннем и начале среднего ордовика, а рифы появились лишь в конце среднего ордовика. Неизмеримо более длительное время оказалось необходимым для начала поздневизейско-серпуховского и пермского циклов рифогенеза. Не исключено, что в этих случаях существовали какие-то иные, дополнительные причины столь длительной задержки начала формирования соответствующих рифов.
Несоответствие временного появления рифостроителей и создаваемых ими рифов отмечены и в случае пермского цикла. Мшанки, одна из важнейших групп, участвовавших в создании рифов, особенно в ранней и средней перми, существовали в карбоне, тубифиты — одни из важных и специфических строителей рифов последнего палеозойского цикла появились еще в позднем карбоне, но настоящее массовое рифостроение началось лишь с начала перми.
Не очень ясна ситуация с началом кембрийского рифообразования. С одной стороны, поскольку рифостроящими каркасообразователями являются только археоциаты, рифы могли формироваться сразу с начала кембрия. С другой не исключено, что становление специфической рифовой экосистемы археоцат и микробиальноводорослевых сообществ, включая эпифитоны, ренальцисы и другие подобные формы, могло сложиться с некоторым запаздыванием, и тогда реализуется ранее предложенный вариант — появление сначала биостромов и биогермов, и лишь затем, с тем или иным запаздыванием, — началось собственно рифообразование [2].
Касаясь вопроса о соотношении времени прекращения рифообразования и границ массовых вымираний, можно констатировать, что также отмечается временное несоответствие прекращения развития рифов и создающих (строящих) их организмов — оно начинается раньше границы биотических кризисов и массовых вымираний, что было отмечено ранее [7—10].
Дело, видимо, в том, что рифовая экосистема в климаксной стадии максимального развития в определенной степени замкнута сама на себя и устойчива при стабильных внешних условиях. При изменении последних она как единая система в трофических и медиотопических отношениях становится неустойчивой, деградирует и распадается, хотя отдельные, составляющие ее таксоны еще существуют. Границы вымираний, «событий», таких как отмеченное выше событие Кельвассер, — это завершающий момент массовых вымираний, вызванных изменениями внешней среды. Подобное обстоятельство фиксируется на границах ордовика и силура, франа-фамена, перми и триаса [5, 7—10].
Менее ясна, как и раньше, ситуация с окончанием кембрийского рифообразования. Не исключено, что очень ограниченный в систематическом отношении биоценоз, где скелетными являлись лишь археоциаты, обусловил прекращение рифостроения одновременно с исчезновением последних.
Таким образом, рифостроящие каркасообразующие организмы появляются в геологической истории раньше, чем начинается образование самих рифов — требуется определенное время для формирования рифового биоценоза, собственной рифовой экосистемы. При изменении внешних условий, которые в итоге приводят к массовым вымираниям, эта рифовая экосистема деградирует и распадается раньше, чем наступает максимум вымирания.
1. Алексеев А.С. Типизация фанерозойских событий массового вымирания организмов // Вестник МГУ. Серия 4. Геология. 2000. № 5. С. 6—14.
2. Кузнецов В.Г. Кембрийское рифообразование в пределах России // Стратиграфия. Геологическая корреляция. 1998. Т. 6. № 6. С. 82—91.
3. Кузнецов В.Г. Палеозойское рифообразование на территории России и смежных стран. М.: ГЕОС, 2000, 228 с.
4. Кузнецов В.Г. Эволюция карбонатонакопления в истории Земли. М.: ГЕОС, 2003. 262 с.
5. Кузнецов В.Г. Рифы позднего ордовика и биологический кризис на границе ордовик-силур // Стратиграфия. Геологическая корреляция. 2018. Т. 26. № 3. С. 24—30. https://doi.org/10.1134/S0869593818030085
6. Кузнецов В.Г., Антошкина А.И. Поздневизейскосерпуховский этап палеозойского рифообразования // Стратиграфия. Геологическая корреляция. 2005. Т. 13. № 4. С. 61—77.
7. Кузнецов В.Г., Журавлева Л.М. Рифовые образования Западно-Канадского бассейна и их нефтегазоносность // Литология и полезные ископаемые. 2018. № 3. С. 257—273. https://doi.org/10.7868/S0024497X18030047
8. Кузнецов В.Г., Журавлева Л.М. Девонское рифообразование в обрамлении Прикаспийской впадины // Литология и полезные ископаемые. 2018. № 5. С. 432—443. https://doi.org/10.1134/S0024497X18050063
9. Кузнецов В.Г. Журавлева Л.М. Рифообразование в эпохи вымираний: граница франа-фамена и девона-карбона // Доклады АН. 2018. Т. 481. № 4. С. 410—413.
10. Кузнецов В.Г., Журавлева Л.М. Геологические и биологические причины прекращения рифообразования, палеозой // Литология и полезные ископаемые. 1919. № 2. С. 119 —129.
11. Кузнецов В.Г., Журавлева Л.М., Лю Шицы. Верхнепермские рифы Южного Китая — последние рифы палеозоя // Литосфера. 2019. Т. 19. № 3. С. 386—392.
12. Рифогенные постройки в палеозое России. М.: Наука, 1997. 167 с.
13. Рифогенные формации и рифы в эволюции биосферы. М.: ПИН, 2011. 228 с.
14. Современные и ископаемые рифы. Термины и определения. Справочник. М.: Недра, 1990. 184 с.
15. Copper P. Evolution, Radiations, and Extinction in Proterozoic to Mid-Paleozoic reefs // The History and Sedimentology of Ancient reef Systems. New York: Kluwer, 2001. P, 89—119.
16. Copper P. Silurian and Devonian reefs: million years of global greenhouse between two Ice ages // SEPM Sp. Publ. 2002. No 72. P. 18—238.
17. Dunham R.J. Classification of carbonate rocks according to deposition texture // Classification of Carbonate rocks. AAPG. Memorr. 1. 19612. P. 108—121.
18. Fluegel E., Fluegel-Kahler E. Phanerozoic reef evolution: Basic Questions and Data Base // Facies. 1992. Vol. 26. P. 167—278.
19. James N.P., Bourque P.A. Reef and mounds // Facies Models: Response to Sea Level Change: Geological association of Canada. 1992. P. 323—347.
20. Kiessling W., Fluegel E. Paleoreefs — a datebase on Phnerozoic Reefs // SEPM Sp. Publ. 2002. No 72. P. 77—92.
21. Kuznetsov V.G. Reef Construction in the Second Half of the Permian and Biotic Crisis at the Permian-Triassic Boundary // Stratigraphy and Geological Correlation. 2008. Vol. 26. No 7. P. 743 —758.
22. Phanerozoic Reef Patterns / W. Kissling, E. Fluegel, J. Golonka, eds. SEPM Sp. Publ. 2002. No 72.
23. The History and Sedimentology of Ancient reef Systems / ed. G.D. Stanly. Kluwer. New York, 2001. 458 p.
24. Webby B. Pattern of Ordovician Reef Developments // SEPM Sp. Publ. No 72. P. 129—179.
доктор геолого-минералогических наук, профессор. Главный научный сотрудник
65, Ленинский проспект, г. Москва 119991, Россия
3, ул. Губкина, г. Москва, 119333, Россия
тел.: +7 (495) 330-39-42
SPIN: 9477-8454
Кузнецов В.Г. АСИНХРОННОСТЬ РАЗВИТИЯ РИФОВ И РИФОСТРОЯЩЕЙ БИОТЫ. ПАЛЕОЗОЙ. Proceedings of Higher Educational Establishments: Geology and Exploration. 2020;1(1):54-62. https://doi.org/10.32454/0016-7762-2020-63-1-54-62
Kuznetsov V.G. ASYNCHRONOUS DEVELOPMENT OF REEFS AND REEF-BUILDING BIOTA. PALEOZOIC. Proceedings of higher educational establishments. Geology and Exploration. 2020;1(1):54-62. (In Russ.) https://doi.org/10.32454/0016-7762-2020-63-1-54-62

23, Миклухо-Маклая ул., Москва 117997, Россия
МГРИ
Телефон: 8 (495) 255-15-10 (доб. 22-89)
E-mail: journal-geology@mgri.ru
E-mail: serovsg@mgri.ru


































