




Содержание
Перейти к:
https://doi.org/10.32454/0016-7762-2025-67-2-100-110
EDN: PRQRPN
Перейти к:
Введение. Брахиоподы эволюционировали на протяжении фанерозоя, и знания о них важны при изучении крупнейших биотических кризисов. Пополнение и ревизия палеонтологической информации требуют регулярного пересмотра оценок влияния кризисов на таксономическое разнообразие данной группы морских организмов.
Цель. Изучение динамики родового разнообразия брахиопод под влиянием наиболее известных массовых вымираний фанерозоя с использованием трех блоков данных, представляющих разные «поколения» палеонтологических компиляций.
Материалы и методы. На основе каждого блока данных построена кривая родового разнообразия брахиопод. При этом три кривые приведены к единой шкале геологического времени. Для массовых вымираний оцениваются снижение числа родов и соотношение с предшествующими и последующими тенденциями изменения разнообразия.
Результаты. Все кривые показывают, что массовые вымирания конца ордовика, рубежей перми и триаса, триаса и юры, мела и палеогена способствовали снижению родового разнообразия брахиопод. Наибольшим было влияние катастрофы на рубеже перми и триаса. Напротив, эффект франско-фаменского (позднедевонского) события был минимальным или отсутствовал вовсе. Тенденции изменения разнообразия до и после кризисных интервалов различались.
Обсуждение результатов. Ретроспективное рассмотрение кривых родового разнообразия брахиопод показывает, что в одних случаях более новая информация говорит о большей интенсивности кризисов, а в других — о меньшей. Более того, смена «поколений» палеонтологических компиляций не способствовала лучшему пониманию динамики числа родов рассматриваемой группы организмов. В этой связи рекомендуется использовать три кривые совместно, а разницу между ними понимать как величину ошибки.
Заключение. Полученные результаты указывают на подверженность брахиопод массовым вымираниям, но не всем из них и в разной степени. Более того, неопределенность ряда оценок сохраняется, что делает актуальным последующие исследования развития брахиопод на критических интервалах геологической истории.
Рубан Д.А. Изменения родового разнообразия брахиопод на интервалах крупнейших массовых вымираний фанерозоя. Proceedings of Higher Educational Establishments: Geology and Exploration. 2025;67(2):100-110. https://doi.org/10.32454/0016-7762-2025-67-2-100-110. EDN: PRQRPN
Ruban D.A. Changes in the generic diversity of brachiopods at the intervals of the largest Phanerozoic mass extinctions. Proceedings of higher educational establishments. Geology and Exploration. 2025;67(2):100-110. (In Russ.) https://doi.org/10.32454/0016-7762-2025-67-2-100-110. EDN: PRQRPN
Брахиоподы являются крупной группой ископаемых организмов, которые эволюционировали на протяжении фанерозоя, а их разнообразие испытывало значительные изменения [15][32][34][67][69]. Изучение этой группы имеет большое значение для понимания биотических катастроф в истории Земли [2][4][10][36][47][48][64], интерес к которым проявляют и отечественные специалисты [1–6]. Известно, что «великие» массовые вымирания [7][35][51] оказали значительное влияние на брахиопод.
Если принять во внимание, что биотические катастрофы могут пониматься как кризисы таксономического разнообразия [40], возникает закономерный вопрос о том, насколько именно оно снижалось. Неизбежная неопределенность ответа на него связана с тем, что накопление, пересмотр и стратиграфическая коррекция палеонтологических данных происходят непрерывно. В этом отношении брахиоподы особенно интересны, так как за четверть века появились три огромных и качественно отличных блока информации о них. Они отражают состояние знаний о стратиграфическом распространении родов брахиопод по состоянию на 1990-е [58], 2000-е [18] и 2010-е годы [42]. Используя их для построения кривых разнообразия, можно не только оценить влияние массовых вымираний на данную группу организмов, но и зафиксировать изменение таких оценок с течением времени.
Целью настоящей статьи является анализ изменений родового разнообразия брахиопод под влиянием пяти «великих» массовых вымираний в свете различных блоков информации. Акцент именно на этих биотических катастрофах связан с высокой степенью их изученности. Более того, интерес к ним изначально формировал основу знаний о массовых вымираниях [51], хотя известны и другие крупные кризисы [29][39][50]. На родовом уровне информация о брахиоподах отличается наибольшей полнотой.
Новизна исследования связана с построением и сравнением кривых, оригинальность которых определяется привязкой к современной версии шкалы геологического времени. Работа имеет не только аналитическую, но и обзорную составляющую, и соответствующая библиографическая информация может представлять ценность для отечественных специалистов. Практическая значимость определяется потребностью в знаниях об изменении таксономического разнообразия стратиграфически важных организмов на событийных уровнях, служащих реперами при геологоразведочных работах, поиске месторождений полезных ископаемых.
Принципы построения и интерпретации кривых разнообразия брахиопод хорошо разработаны в современной науке [27][36][44][61][64]. В настоящей работе учитывается количество родов данной группы организмов в веках всех периодов фанерозоя. Соответствующие кривые выравниваются по абсолютной шкале геологического времени.
Были использованы три источника информации [18][42][58]. Данные о родовом разнообразии брахиопод из работы [58] извлекались в помощью онлайн-инструмента, доступного по ссылке: https://strata.geology.wisc.edu/jack/. В сводке [18] данные даются в текстовом и графическом видах. База данных [42] содержит специальную форму для извлечения требующейся информации. Принципиально важно то, что вышеуказанные источники используют разные шкалы геологического времени, в том числе с региональными и устаревшими подразделениями. В этой связи потребовалась привязка стратиграфической основы каждого источника к современной версии международной шкалы с установленными для границ веков абсолютными отметками [24] (см. обновления этой шкалы на https://stratigraphy.org/).
Извлечение информации из вышеуказанных источников и стратиграфическая коррекция позволили построить кривые COM (от слова «compendium», определяющего тип источника; данные [58]), TRE (от слова «treatise», относящегося к серии источника; данные [18]), PDB (от сокращенного названия базы данных «Paleobiology Database»; данные [42]). На каждую из них проецировались временные отметки пяти «великих» массовых вымираний, имевших место в конце ордовика, в позднем девоне, на рубежах перми и триаса, триаса и юры, мела и палеогена. Далее определялась их выраженность на каждой кривой, т.е. характер изменений последней на интервале массового вымирания, а также до и после него.
Массовое вымирание конца ордовика оказало исключительно сильное воздействие на морские экосистемы [26][60][65]. Подверженность ему брахиопод хорошо известна [9][19][23][30][53]. Все три кривые фиксируют снижение родового разнообразия на интервале данного массового вымирания (рис. 1). Кривая COM показывает сокращение числа родов в 1,2 раза. Оно последовало за локальным пиком в первой половине позднего ордовика и продолжилось в раннем силуре. Кривая TRE обнаруживает менее выраженный эффект (снижение родового разнообразия в <1,1 раза), который проявился на фоне долговременной негативной тенденции. Интересно отметить, что самый сильный спад числа родов имел место не в конце ордовика (см. выше), а в начале силура, после чего разнообразие стало активно восстанавливаться. Кривая PDB говорит о сокращении разнообразия в 2,5 раза, которое имело место непосредственно в конце ордовика. Оно произошло сразу после пика в середине позднего ордовика и замедлилось на непродолжительное время в раннем силуре, после чего стало постепенно расти.
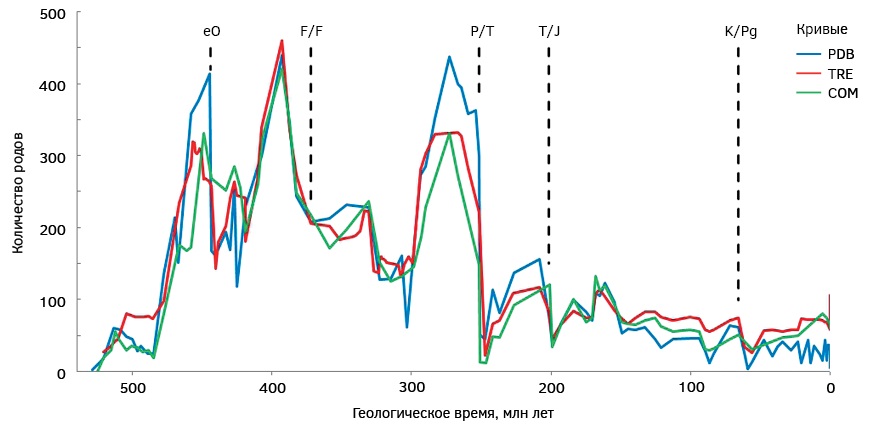
Рис. 1. Динамика родового разнообразия брахиопод в фанерозое: COM — по данным [58], TRE — по данным [18], PDB — по данным [42]. Массовые вымирания: eO — конца ордовика, F/F — франско-фаменское (позднедевонское), P/T — рубежа перми и триаса, T/J — перехода от триаса к юре, K/Pg — рубежа мела и палеогена
Fig. 1. Dynamics of the generic diversity of brachiopods in the Phanerozoic: COM — according to [58], TRE — according to [18], PDB — according to [42]. Mass extinctions: eO — end-Ordovician, F/F — Frasnian-Famennain (Late Devonian), P/T — Permian-Triassic boundary, T/J — Triassic-Jurassic transition, K/Pg — Cretaceous-Paleogene boundary
Франско-фаменское (позднедевонское) массовое вымирание стало одной из самых значимых палеозойских биотических катастроф [3][6][20][49][55]. Известно, что оно сказалось на брахиоподах [2][14][17], что фиксируется не всеми кривыми их родового разнообразия (рис. 1). Кривая COM показывает сокращение числа таксонов в 1,3 раза. При этом соответствующая тенденция установилась еще с начала среднего девона, а отмеченное сокращение стало ее кульминацией. Кривая TRE выявляет незначительное (<1,1 раза) сокращение числа родов, которое последовало за значительно более сильным спадом разнообразия в предшествующие века. В самом начале карбона число родов уменьшилось сильнее. Кривая PDB указывает на небольшое (<1,1 раза) увеличение разнообразия в фаменском веке в сравнении с франским. Как и в предыдущих случаях, этому предшествовал длительный спад числа родов, а зафиксированный на интервале рассматриваемого массового вымирания слабый рост сохранялся и в начале карбона.
Массовое вымирание на рубеже перми и триаса, вполне вероятно, является самой сильной биотической катастрофой фанерозоя, оказавшей беспрецедентное воздействие на морские экосистемы [12][25][43]. Его опустошительное влияние на брахиопод уже отмечалось исследователями [28][52], хотя некоторые представители данной группы организмов пережили его [16]. Негативные эффекты данного события фиксируются всеми кривыми (рис. 1). Кривая COM выявляет очень интенсивное снижение числа родов (в 11,4 раза). Оно наложилось на тенденцию к снижению разнообразия, установившуюся еще в середине перми и достигшую кульминации в оленекском веке раннего триаса. Кривая TRE также показывает очень сильное сокращение числа родов (в 9,6 раза), который стал кульминацией реализовывавшейся ранее тенденции. Кривая PDB указывает на менее интенсивное, но все равно значительное (в 5,8 раза) снижение таксономического разнообразия. Последнее началось еще в середине перми, прервалось кратким эпизодом незначительного роста, после чего усилилось на интервале массового вымирания, продолжившись в раннем триасе.
На переходе от триаса к юре имело место еще одно массовое вымирание, которое отличалось комплексностью [13][41][54][56][66]. Брахиоподы оказались подвержены влиянию этого события [21][63]. Кривые их таксономического разнообразия фиксируют отрицательный пик вблизи рубежа триаса и юры (рис. 1). Кривая COM показывает четко выраженное сокращение числа родов (в 3,6 раза). Оно прервало ранее имевшее место увеличение разнообразия, которое возобновилось в первой половине ранней юры. Кривая TRE указывает на менее интенсивный кризис, когда число родов снизилось в 1,8 раза. Ему предшествовал заметный спад в рэтском веке, однако в ранней юре начался новый рост. Кривая PDB позволяет увидеть аналогичное сокращение разнообразия (также в 1,8 раза), которое продолжило установившуюся незадолго до этого тенденцию. Как и в двух других случаях, увеличение числа родов началось в ранней юре.
Массовое вымирание на рубеже мела и палеогена является самым известным и при этом оказавшим сильное воздействие на морскую биоту [5][8][31][37]. Из-за общего сокращения таксономического разнообразия брахиопод во второй половине мезозоя — кайнозое влиянию этой катастрофы на них уделялось не столь много внимания, хотя отдельные исследования фиксировали соответствующие эффекты [33][57][62]. Иногда результаты оказывались противоречивыми [38]. Тем не менее все построенные кривые довольно уверенно фиксируют воздействие этого массового вымирания на рассматриваемую группу организмов (рис. 1). Кривая COM показывает снижение числа родов в 1,6 раза. Оно имело место сразу после небольшого положительного пика в маастрихте, и за ним последовал медленный, но долгий рост. Кривая TRE указывает на больший (в 2,1 раза) спад разнообразия, который продолжился в палеоцене. При этом во второй половине мела (сантонский — маастрихтский века) число таксонов медленно, но направленно росло. Кривая PDB отображает еще более интенсивный (в 2,3 раза) спад родового разнообразия. Тенденция к сокращению числа таксонов установилась еще в маастрихтском веке и достигла кульминации в зеландском веке, после чего разнообразие так и не вернулось к значениям конца мезозоя.
Рассмотренные кривые родового разнообразия брахиопод различаются качественно, так как основаны на блоках информации [18][42][58], скомпилированной разными способами. В этой связи имеет смысл обобщить полученные результаты. Получается, что из пяти «великих» массовых вымираний четыре уверенно фиксируются палеонтологическими данными, так как им соответствуют негативные эффекты, отображаемые всеми кривыми. Исключением оказывается позднедевонская катастрофа, значительное влияние которой на брахиопод устанавливается только одной и при этом самой «старой» кривой. Это означает, что или рассматриваемая группа организмов оказалось устойчивой к действию данного массового вымирания в глобальном масштабе, или наши знания об эволюции позднедевонских брахиопод отличаются неполнотой. Также эффект этой биотической катастрофы мог носить кратковременный характер, а потому не фиксируется при вековом разрешении кривых.
Еще одно заслуживающее внимания наблюдение касается выражения негативных эффектов массовых вымираний на кривых родового разнообразия брахиопод. Как следует из представленных выше результатов, однозначными выглядят только две ситуации, а именно установление тенденции к сокращению разнообразия до катастрофы на рубеже перми и триаса и диверсификация сразу после вымирания на рубеже триаса и юры. В остальных случаях между кривыми фиксируется разница. С учетом вышесказанного получаем, что влияние рассматриваемых биотических катастроф на брахиопод видно четче, чем их соотношение с предшествующими и последующими тенденциями изменения разнообразия.
Хотя все три кривые сохраняют свою актуальность, нельзя игнорировать тот факт, что между появлением соответствующих источников информации [18][42][58] проходило некоторое время, и эти источники представляют разные «поколения» палеонтологических компиляций. Смена этих «поколений» связана с накоплением и ревизией информации о брахиоподах, а также видоизменением шкалы геологического времени (в частности, на некоторых интервалах она становилась более дробной, а на других — менее дробной). В таком случае полученные результаты (рис. 1) могут сравниваться в ретроспективном отношении. Совершенствование палеонтологической и стратиграфической информации в течение трех десятилетий позволило обнаружить более интенсивное и более резкое воздействие биотического кризиса конца ордовика на родовое разнообразие брахиопод, полное отсутствие влияния франско-фаменского кризиса, менее интенсивное и нерезкое проявление кризиса на рубеже перми и триаса, менее интенсивный и менее резкий стресс на рубеже триаса и юры, а также более интенсивное, но менее резкое воздействие кризиса на рубеже мела и палеогена. В указанных случаях интенсивность означает величину сокращения числа родов (в разы), а резкость — нарушение предшествовавшей тенденции (наибольшая резкость имеет место в том случае, если вызываемый катастрофой спад разнообразия нарушает ранее имевший место долговременный рост). Таким образом оказывается, что прогресс палеонтологических и стратиграфических знаний по-разному влияет на понимание изменений родового разнообразия брахиопод на интервалах массовых вымираний. Одни из них оказываются более интенсивными и/или резкими, чем считалось ранее, а другие — менее интенсивными и/или резкими.
Общий вид построенных кривых (рис. 1) поднимает и еще один вопрос. Казалось бы, продолжающееся накопление палеонтологической информации должно выражаться в более высоких оценках разнообразия групп ископаемых организмов с течением времени. Даже если отдельные таксоны признаются невалидными или объединяются, то параллельно с ними обнаруживаются все новые и новые. Однако это предположение оказывается полностью несостоятельным. Новейшая кривая PDB лишь на отдельных участках проходит выше более «старых» кривых COM и TRE, а кривая COM кое-где занимает максимальную позицию по отношении к двум другим (рис. 1). Конечно, это можно объяснить изменениями дробности шкалы геологического времени, но лишь отчасти. Наиболее интересные примеры обнаруживаются в мезозое, где стратиграфическая основа использованных источников палеонтологической информации большей частью одинакова [18][42][58], но при этом новейшая кривая PDB проходит часто ниже двух других кривых. Компиляция информации о брахиоподах на более современной основе, а именно с использованием постоянно пополняемой базы данных [42], не позволила достичь уровня знаний, который обеспечивался традиционными способами работы [18][58]. Этот вывод также подтверждает мысль о том, что три рассмотренные кривые лучше всего использовать совместно, не отдавая явного предпочтения наиболее «свежей» из них. В таком случае различия между кривыми указывают на приблизительный размер ошибки, который надо учитывать при интерпретациях. Дальнейший прогресс может быть связан с созданием палеонтологических баз данных с применением технологий искусственного интеллекта [45][59][68]. При этом важно понимать, что некоторые биотические события известны еще с XIX века [11][22][46], и, следовательно, их полная нивелировка вряд ли ожидаема.
Сравнивая пять «великих» массовых вымираний по их воздействию на родовое разнообразие брахиопод и при этом принимая равнозначность построенных кривых, можно предполагать, что наибольший стресс данной группы организмов был связан с катастрофой на рубеже перми и триаса, а наименьший — с позднедевонским кризисом. Вторым по степени влияния оказывается вымирание рубежа триаса и юры, третьим — рубежа мела и палеогена, четвертым — конца ордовика.
Проведенное исследование позволило сделать следующие выводы. Во-первых, построение и сравнение кривых родового разнообразия брахиопод позволило подтвердить их подверженность влиянию четырех из пяти «великих» массовых вымираний. Во-вторых, результаты выявляют значительную неопределенность в понимании воздействия франско-фаменской биотической катастрофы на рассматриваемую группу организмов. В-третьих, смена «поколений» палеонтологических компиляций привела к тому, что влияние один массовых вымираний оказалось сильнее, а других — слабее, и в целом она не способствовала лучшему пониманию их эффектов.
В практическом отношении полученные результаты указывают на необходимость более тщательной работы с палеонтологической и стратиграфической информацией при определении событийных уровней, значимых для геологоразведочных работ. Отдельные группы организмов могут оказаться более или менее полезными при установлении и корреляции маркирующих горизонтов, связанных с интервалами массовых вымираний. Перспективным направлением для последующих исследований является разработка более совершенных инструментов компиляции и интерпретации палеонтологической информации, которые бы позволяли полноценно учитывать ранее накопленные данные без стратиграфических неточностей.
Рубан Д.А. — разработал концепцию статьи, проанализировал палеонтологическую информацию, интерпретировал полученные результаты, подготовил текст статьи, окончательно утвердил публикуемую версию статьи и согласен принять на себя ответственность за все аспекты работы.
Dmitry A. Ruban — developed the article concept, analyzed paleontological information, interpreted results, prepared the article text, approved the final version of the article and accepted the responsibility for all aspects of the work.
1. Алексеев А.С. Типизация фанерозойских событий массового вымирания организмов. Вестник Московского университета. Серия 4: Геология. 2000. № 5. С. 6—14.
2. Афанасьева Г.А. Разнообразие и распространение брахиопод отряда Chonetida в карбоне. Палеонтологический журнал. 2022. № 5. С. 19—28.
3. Бараш М.С. Причины великого массового вымирания морских организмов в позднем девоне. Океанология. 2016. № 6. С. 946—958.
4. Грунт Т.А. Таксономия и основные направления развития замковых брахиопод отряда Athyridida. Ученые записки Казанского университета. Естественные науки. 2010. Т. 152. Кн. 4. С. 123—134.
5. Кочергин Д.В., Грановская Н.В. Признаки катастрофического космического события на границе меловых и палеогеновых отложений Северо-Западного Кавказа. Грозненский естественнонаучный бюллетень. 2024. № 1. С. 42—48.
6. Кузнецов В.Г., Журавлева Л.М. Рифообразование в эпохи массовых вымираний: граница франа — фамена и девона — карбона. Доклады Академии наук. 2018. № 4. С. 410—413.
7. Algeo T.J., Shen J. Theory and classification of mass extinction causation. National Science Review. 2024. V. 11. P. nwad237.
8. Alvarez L.W., Alvarez W., Asaro F., Michel H.V. Extraterrestrial cause for the Cretaceous-Tertiary extinction. Science. 1980. V. 208. P. 1095—1108.
9. Baarli B.G., Huang B., Johnson M.E. The deep-water, high-diversity Edgewood-Cathay brachiopod Fauna and its Hirnantian counterpart. Palaeogeography, Palaeoclimatology, Palaeoecology 2024. V. 642. P. 112153.
10. Baeza-Carratalá J.F., García Joral F. The last representatives of the Superfamily Wellerelloidea (Brachiopoda, Rhynchonellida) in the westernmost Tethys (Iberian paleomargins) prior to their demise in the early Toarcian Mass Extinction Event. Journal of Paleontology. 2022. V. 96. P. 991—1023.
11. Bailey R. Mass extinctions and their causes. Geology Today. 2024. V. 40. P. 21—28.
12. Benton M.J., Twitchett R.J. How to kill (almost) all life: The end-Permian extinction event. Trends in Ecology and Evolution. 2003. V. 18. P. 358—365.
13. Bond A.D., Dickson A.J., Ruhl M., Bos R., van de Schootbrugge B. Globally limited but severe shallow-shelf euxinia during the end-Triassic extinction. Nature Geoscience. 2023. V. 16. P. 1181—1187.
14. Brisson S.K., Pier J.Q., Beard J.A., Fernandes A.M., Bush A.M. Niche conservatism and ecological change during the Late Devonian mass extinction. Proceedings of the Royal Society B: Biological Sciences. 2023. V. 290. P. 20222524.
15. Carlson S.J. The Evolution of Brachiopoda. Annual Review of Earth and Planetary Sciences. 2016. V. 44. P. 409—438.
16. Chen Z.-Q., Kaiho K., George A.D. Survival strategies of brachiopod faunas from the end-Permian mass extinction. Palaeogeography, Palaeoclimatology, Palaeoecology. 2005. V. 224. P. 232—269.
17. Copper P. Evaluating the Frasnian-Famennian mass extinction: comparing brachiopod faunas. Acta Palaeontologica Polonica. 1998. V. 43. P. 137—154.
18. Curry G.B., Brunton C.H.C. Stratigraphic distribution of brachiopods. Treatise on Invertebrate Paleontology. Part H. Brachiopoda. Revised. Vol. 6. Boulder: GSA, 2007. P. 2901—3081.
19. Darroch S.A.F., Wagner P.J. Response of beta diversity to pulses of Ordovician-Silurian mass extinction. Ecology. 2015. V. 96. P. 532—549.
20. De Vleeschouwer D., Da Silva A.-C., Sinnesael M., Chen D., Day J.E., Whalen M.T., Guo Z., Claeys P. Timing and pacing of the Late Devonian mass extinction event regulated by eccentricity and obliquity. Nature Communications. 2017. V. 8. P. 2268.
21. Deng S., Lu Y., Xu D. Progress and review of the studies on the end-Triassic mass extinction event. Science in China, Series D: Earth Sciences. 2005. V. 48. P. 2049—2060.
22. Eldredge N. Revisiting Clarence King’s “Catastrophism and Evolution” (1877). Biological Theory. 2019. V. 14. P. 247—253.
23. Finnegan S., Rasmussen C.M.Ø., Harper D.A.T. Identifying the most surprising victims of mass extinction events: An example using Late Ordovician brachiopods. Biology Letters. 2017. V. 13. P. 20170400.
24. Gradstein F.M., Ogg J.G., Schmitz M., Ogg G. (Editors). Geologic Time Scale 2020. Amsterdam: Elsevier, 2020. 1390 p.
25. Grasby S.E., Ardakani O.H., Liu X., Bond D.P.G., Wignall P.B., Strachan L.J. Marine snowstorm during the Permian-Triassic mass extinction. Geology. 2024. V. 52. P. 120—124.
26. Harper D.A.T. Late Ordovician Mass Extinction: Earth, fire and ice. National Science Review. 2024. V. 11. P. nwad319.
27. Harper D.A.T., Drachen A. The Orthida: The rise and fall of a great Palaeozoic brachiopod clade. Special Papers in Palaeontology. 2010. V. 84. P. 107—117.
28. He W.-H., Shi G.R., Twitchett R.J., Zhang Y., Song H.-J., Yue M.-L., Wu S.-B., Wu H.-T., Yang T.-L., Xiao Y.-F. Late Permian marine ecosystem collapse began in deeper waters: Evidence from brachiopod diversity and body size changes. Geobiology. 2015. V. 13. P. 123—138.
29. House M.R. Strength, timing, setting and cause of mid-Palaeozoic extinctions. Palaeogeography, Palaeoclimatology, Palaeoecology. 2002. V. 181. P. 5—25.
30. Huang B., Rong J., Cocks L.R.M. Global palaeobiogeographical patterns in brachiopods from survival to recovery after the end-Ordovician mass extinction. Palaeogeography, Palaeoclimatology, Palaeoecology. 2012. V. 317—318. P. 196—205.
31. Jablonski D., Raup D.M. Selectivity of end-Cretaceous marine bivalve extinctions. Science. 1995. V. 268. P. 389—391.
32. Jain S. Fundamentals of Invertebrate Palaeontology. Macrofossils. New Delhi: Springer, 2016. 405 p.
33. Johansen M.B. Background extinction and mass extinction of the brachiopods from the chalk of northwest Europe. Palaios. 1989. V. 4. P. 243—250.
34. Jones R.W. Applied Palaeontology. Cambridge: Cambridge University Press, 2012. 434 p.
35. Kaiho K. Relationship between extinction magnitude and climate change during major marine and terrestrial animal crises. Biogeosciences. 2022. V. 19. P. 3369—3380.
36. Liang Y., Strotz L.C., Topper T.P., Holmer E.L., Budd G.E., Chen Y., Fang R., Hu Y., Zhang Z. Evolutionary contingency in lingulid brachiopods across mass extinctions. Current Biology. 2023. V. 33. P. 1565—1572.
37. Lowery C.M., Bralower T.J., Owens J.D., Rodriguez-Tovar F.J., Jones H., Smit J., Whalen M.T., Claeys P., Farley K., Gulick S.P.S., Morgan J.V., Green S., Chenot E., Christeson G.L., Cockell C.S., Coolen M.J.L., Ferriere L., Gebhardt C., Goto K., Kring D.A., Lofi J., Ocampo-Torres R., Perez-Cruz L., Pickersgill A.E., Poelchau M.H., Rae A.S.P., Rasmussen C., Rebolledo-Vieyra M., Riller U., Sato H., Tikoo S.M., Tomioka N., Urrutia-Fucugauchi J., Vellekoop J., Wittman A., Xiao L., Yamaguchi K.E., Zylberman W. Rapid recovery of life at ground zero of the end-Cretaceous mass extinction. Nature. 2018. V. 558. P. 288—291.
38. Macleod N., Rawson P.F., Forey P.L., Banner F.T., Boudagher-Fadel M.K., Bown P.R., Burnett J.A., Chambers P., Culver S., Evans S.E., Jeffry C., Kaminski M.A., Lord A.R., Milner A.C., Milner A.R., Morris N., Owen E., Rosen B.R., Smith A.B., Taylor P.D., Urquhart E., Young J.R. The Cretaceous-Tertiary biotic transition. Journal of the Geological Society. 1997. V. 154. P. 265—292.
39. McGhee G.R., Clapham M.E., Sheehan P.M., Bottjer D.J., Droser M.L. A new ecological-severity ranking of major Phanerozoic biodiversity crises. Palaeogeography, Palaeoclimatology, Palaeoecology. 2013. V. 370. P. 260—270.
40. Padian K. Measuring and comparing extinction events: Reconsidering diversity crises and concepts. Integrative and Comparative Biology. 2018. V. 58. P. 1191—1203.
41. Pálfy J., Mortensen J.K., Carter E.S., Smith P.L., Friedman R.M., Tipper H.W. Timing the end-Triassic mass extinction: First on land, then in the sea?. Geology. 2000. V. 28. P. 39—42.
42. Paleobiology Database, 2024. URL: https://paleobiodb.org/ (дата обращения: 24.04.2024).
43. Payne J.L., Clapham M.E. End-Permian mass extinction in the oceans: An ancient analog for the twenty-first century? Annual Review of Earth and Planetary Sciences. 2012. V. 40. P. 89—111.
44. Penny A., Kröger B. Impacts of spatial and environmental differentiation on early Palaeozoic marine biodiversity. Nature Ecology and Evolution. 2019. V. 3. P. 1655—1660.
45. Peters S.E., Zhang C., Livny M., Ré C. A machine reading system for assembling synthetic paleontological databases. PLoS ONE. 2014. V. 9. P. e113523.
46. Posenato R. The end-Permian mass extinction (EPME) and the early triassic biotic recovery in the western Dolomites (Italy): state of the art. Bollettino della Societa Paleontologica Italiana. 2019. V. 58. P. 11—34.
47. Powell M.G. Timing and selectivity of the Late Mississippian mass extinction of brachiopod genera from the Central Appalachian Basin. Palaios. 2008. V. 23. P. 525—534.
48. Qiao L., Qie W. Palaeobiogeographic dynamics of brachiopod faunas during the Frasnian-Famennian biotic crisis in South China. Palaeobiodiversity and Palaeoenvironments. 2019. V. 99. P. 91—99.
49. Racki G. A volcanic scenario for the Frasnian– Famennian major biotic crisis and other Late Devonian global changes: More answers than questions? Global and Planetary Change. 2020. V. 189. P. 103174.
50. Rampino M.R., Caldeira K., Rodriguez S. Sixteen mass extinctions of the past 541 My correlated with 15 pulses of Large Igneous Province (LIP) volcanism and the 4 largest extraterrestrial impacts. Global and Planetary Change. 2024. V. 234. P. 104369.
51. Raup D.M., Sepkoski J.J. Mass extinctions in the marine fossil record. Science. 1982. V. 215. P. 1501—1503.
52. Rong J.-Y., Shen S.-Z. Comparative analysis of the end-Permian and end-Ordovician brachiopod mass extinctions and survivals in South China. Palaeogeography, Palaeoclimatology, Palaeoecology. 2002. V. 188. P. 25—38.
53. Rong J., Harper D.A.T., Huang B., Li R., Zhang X., Chen D. The latest Ordovician Hirnantian brachiopod faunas: New global insights. Earth-Science Reviews. 2020. V. 208. P. 103280.
54. Ruban D.A. A review of the Late Triassic conodont conundrum: survival beyond biotic perturbations. Palaeobiodiversity and Palaeoenvironments. 2022. V. 102. P. 373—382.
55. Sandberg C.A., Morrow J.R., Ziegler W. Late Devonian sea-level changes, catastrophic events, and mass extinctions. Special Paper of the Geological Society of America. 2002. V. 356. P. 473—487.
56. Schoepfer S.D., Shen J., Sano H., Algeo T.J. Onset of environmental disturbances in the Panthalassic Ocean over one million years prior to the Triassic-Jurassic boundary mass extinction. Earth-Science Reviews. 2002. V. 224. P. 103870.
57. Schrøder A.E., Surlyk F. Adaptive brachiopod morphologies in four key environments of the Late Cretaceous–Danian Chalk Sea of northern Europe: A comparative study. Cretaceous Research. 2020. V. 107. P. 104288.
58. Sepkoski J.J. A compendium of fossil marine animal genera. Bulletins of American Paleontology. 2002. V. 363. P. 1—560.
59. Serra F., Balseiro D., Monnet C., Randolfe E., Bignon A., Rustan J.J., Bault V., Munoz D.F., Vaccari N.E., Martinetto M., Crônier C., Waisfeld B.G. A dynamic and collaborative database for morphogeometric information of trilobites. Scientific Data. 2023. V. 10. P. 841.
60. Sheehan P.M. The late Ordovician mass-extinction. Annual Review of Earth and Planetary Sciences. 2001. V. 29. P. 331—364.
61. Shen S-Z., Shi G.R. Paleobiogeographical extinction patterns of Permian brachiopods in the Asian-western Pacific region. Paleobiology. 2002. V. 28. P. 449—463.
62. Surlyk F., Johansen M.B. End-Cretaceous brachiopod extinctions in the Chalk of Denmark. Science. 1984. V. 223. P. 1177—1179.
63. Tomašových A., Siblík M. Evaluating compositional turnover of brachiopod communities during the end-Triassic mass extinction (Northern Calcareous Alps): Removal of dominant groups, recovery and community reassembly. Palaeogeography, Palaeoclimatology, Palaeoecology. 2007. V. 244. P. 170—200.
64. Vörös A., Kocsis Á.T., Pálfy J. Mass extinctions and clade extinctions in the history of brachiopods: Brief review and a post-Paleozoic case study. Rivista Italiana di Paleontologia e Stratigrafia. 2019. V. 125. P. 711—724.
65. Wang G., Zhan R., Percival I.G. The end-Ordovician mass extinction: A single-pulse event?. Earth-Science Reviews. 2019. V. 192. P. 15—33.
66. Wignall P.B., Atkinson J.W. A two-phase end-Triassic mass extinction. Earth-Science Reviews. 2020. V. 208. P. 103282.
67. Williams A., Hurst J.M. Brachiopod evolution. Developments in Palaeontology and Stratigraphy. 1977. V. 5. P. 79—121.
68. Zhang C., Ré C., Cafarella M., De Sa C., Ratner A., Shin J., Wang F., Wu S. DeepDive: Declarative knowledge base construction. Communications of the ACM. 2017. V. 60. P. 93—102.
69. Zhang Z., Augustin M., Payne J.L. Phanerozoic trends in brachiopod body size from synoptic data. Paleobiology. 2015. V. 41. P. 491—501.
Рубан Дмитрий Александрович — Philosophiae Doctor (Университет Претории, ЮАР), кандидат геолого-минералогических наук, доцент, доцент ЮФУ.
43, ул. 23-я линия, Ростов-на-Дону 344019
тел.: +7 (903) 463-43-44
Нет
Рубан Д.А. Изменения родового разнообразия брахиопод на интервалах крупнейших массовых вымираний фанерозоя. Proceedings of Higher Educational Establishments: Geology and Exploration. 2025;67(2):100-110. https://doi.org/10.32454/0016-7762-2025-67-2-100-110. EDN: PRQRPN
Ruban D.A. Changes in the generic diversity of brachiopods at the intervals of the largest Phanerozoic mass extinctions. Proceedings of higher educational establishments. Geology and Exploration. 2025;67(2):100-110. (In Russ.) https://doi.org/10.32454/0016-7762-2025-67-2-100-110. EDN: PRQRPN

23, Миклухо-Маклая ул., Москва 117997, Россия
МГРИ
Телефон: 8 (495) 255-15-10 (доб. 22-89)
E-mail: journal-geology@mgri.ru
E-mail: serovsg@mgri.ru


































